Abstract
The demand for recombinant proteins for analytic and therapeutic purposes is increasing; however, most currently used bacterial production systems accumulate the recombinant proteins in the intracellular space, which requires denaturating procedures for harvesting and functional testing. We here present a novel FimH-based expression system that enables display of fully functional eukaryotic proteins while preventing technical difficulties in translocating, folding, stabilizing and isolating the displayed proteins. As examples, Gaussia Luciferase (GLuc), epidermal growth factor (EGF), transforming growth factor-α (TGF-α) and epiregulin (EPRG) were expressed as FimH fusion proteins on the surface of E. coli bacteria.
The fusion proteins were functionally active and could be released from the bacterial surface by specific proteolytic cleavage into the culture supernatant allowing harvesting of the produced proteins. EGFR ligands, produced as FimH fusion proteins and released by proteolytic cleavage, bound to the EGF receptor (EGFR) on cancer cells inducing EGFR phosphorylation.
In another application of the technology, GLuc-FimH expressed on the surface of bacteria was used to track tumor-infiltrating bacteria by bioluminescence imaging upon application to mice, thereby visualizing the colonization of transplanted tumors. The examples indicate that the FimH-fusion protein technology can be used in various applications that require functionally active proteins to be displayed on bacterial surfaces or released into the culture supernatant.
fimh antibody cusabio
Introduction
Bacterial surface display of recombinant proteins has become an attractive strategy for a broad range of applications such as production of bioadsorbents1, generation of cellular biosensors, development of novel vaccine platforms3, screening of antibody libraries4 and whole-cell biocatalysis.
Generally, the procedure requires the fusion of the protein-of-interest (POI) to a bacterial surface protein to display the POI on the surface of the genetically modified bacteria. Several surface-anchoring motifs like LPP-OmpA, LamB, PhoE, ice nucleation protein (INP) and auto-transporter are employed as carrier proteins for crossing the bacteria membrane.
- Despite the successful approaches, several problems remain to be solved, including the substantially reduced functional activity of the displayed proteins. Compared with their soluble form, surface-anchored β-lactamase fused to the translation unit (TU) of an auto-transporter shows substantially reduced catalytic activities. A similar experience was made when displaying sorbitol dehydrogenase.
- A major problem in the use of auto-transporters arises from the tertiary structure of the passenger domains and the size of the central cavity that permits translocating only small proteins. It seems not only to be a matter of size since even the 62 amino acids protein aprotinin is not efficiently translocated through the outer membrane9.
- Translocation by auto-transporters is very sensitive to structure of the passenger proteins that consist of a β-strands backbone with at least 300 amino acids thereby substantially limiting the applicability to variety of potential cargos10.
- As an alternative approach, protein sequences derived from the major E. coli lipoprotein (Lpp) were fused to the N-terminus of the POI to direct the protein to the outer membrane11,12. The system consists of two key anchoring motifs; the Lpp-derived signal sequence at the N-terminus to target the fusion protein to the inner surface of the outer membrane, and the outer membrane protein A (OmpA)-derived transmembrane region to transfer the protein across the outer membrane12.
- Since its introduction by Ghrayeb and Inouye13 in 1984, the Lpp-OmpA display method is facing difficulties including the low expression rate and the insufficient translocation efficiency which may be due to steric hindrance and incorrect folding when anchoring in the outer membrane14,15.
- In Gram-negative bacteria the outer membrane generally acts as a barrier to restrict the protein export from the cell interior; only pilins, flagellins, specific surface enzymes, and a few bacterial toxins are transported across the outer membrane16.
- These natural display systems have the benefit of being optimized for transporting and folding protein units to build polymeric structures on the extracellular surface making the display system attractive for biotechnological applications. We here used the fimbriae protein FimH, the mannose-specific adhesin of the E. coli type-1 fimbriae, for the extracellular display of recombinant proteins.
- Type-1 fimbriae are composed of up to 3,000 copies of the subunit FimA, that form the pilus rod, as well as the subunits FimF, FimG and FimH building the distal tip fibrillum17,18. In initial studies, Pallesen and colleagues used the positions 225 and 258 within the FimH adhesin to display the preS2 domain of the hepatitis B surface antigen or an epitope from cholera toxin19.
- Both positions within the FimH protein proved to be suitable for the integration of peptides of up to 56 amino acids which could be produced, displayed on the cell surface and partially conserved the adhesive function of FimH19.
- Longer peptide or full length proteins displayed by FimH in that position were so far not reported. While short polypeptides used for vaccines could be displayed, the technique failed in functionally expressing large proteins like enzymes or cytokines.
- Here we identified alternative positions within the FimH protein to display larger proteins in a functionally active fashion. Based on the 3D modelling of E. coli type-1 pili20 we identified the N-terminus of the FimH domain on the fimbriae tip as a suitable integration site of a larger protein.
- As examples, we genetically linked Gaussia luciferase (GLuc) and human epidermal growth factor (EGF), tumor growth factor-a (TGF-α) and epiregulin (EREG), all ligands of the epidermal growth factor receptor (EGFR), to FimH.
- Expressed by transformed E. coli, the proteins conserved their functional capacities. In particular, GLuc-FimH displaying E. coli bacteria were tracked during colonization of syngeneic tumors in an immunocompetent mouse model of pancreatic cancer during a six week period without losing GLuc activity.
- Bacteria with surface displayed proteins can be used for screening purposes and, furthermore, can be released in a functionally active form by specific proteolytic cleavage making the strategy attractive for protein production without the need to disrupt the bacteria by harsh procedures.
) Recombinant Escherichia coli protein FimH(fimH) Recombinant Escherichia coli protein FimH(fimH) |
| AP71165 | SAB | 1mg | EUR 2826 |
|
| Recombinant Escherichia coli protein FimH(fimH) |
| AP72985 | SAB | 1mg | EUR 3389 |
|
) Recombinant Escherichia coli Protein FimH(fimH) Recombinant Escherichia coli Protein FimH(fimH) |
| AP76639 | SAB | each | Ask for price |
|
 Antibody) Type 1 fimbrin D-mannose specific adhesin (fimH) Antibody Type 1 fimbrin D-mannose specific adhesin (fimH) Antibody |
| abx345775-100g | Abbexa | 100 µg | Ask for price |
| Type 1 fimbrin D-mannose specific adhesin (fimH) Antibody |
| abx345775-20g | Abbexa | 20 µg | EUR 250 |
| Type 1 fimbrin D-mannose specific adhesin (fimH) Antibody |
| abx345775-50g | Abbexa | 50 µg | EUR 350 |
) Recombinant Escherichia coli FimH Protein (aa 22-300) Recombinant Escherichia coli FimH Protein (aa 22-300) |
| VAng-Lsx02377-1mgEcoli | Creative Biolabs | 1 mg (E. coli) | EUR 5751.6 |
Description: Escherichia coli protein FimH, recombinant protein. |
| Recombinant Escherichia coli FimH Protein (aa 22-300) |
| VAng-Lsx02377-500gEcoli | Creative Biolabs | 500 µg (E. coli) | EUR 3657.6 |
Description: Escherichia coli protein FimH, recombinant protein. |
| Recombinant Escherichia coli FimH Protein (aa 22-300) |
| VAng-Lsx02377-50gEcoli | Creative Biolabs | 50 µg (E. coli) | EUR 1083.6 |
Description: Escherichia coli protein FimH, recombinant protein. |
) OPCA04413-20UG - FIMH Recombinant Protein (Escherichia coli) OPCA04413-20UG - FIMH Recombinant Protein (Escherichia coli) |
| OPCA04413-20UG | Aviva Systems Biology | 20ug | EUR 620 |
- FIMH Recombinant Protein (Escherichia coli)
|
) OPCA04413-100UG - FIMH Recombinant Protein (Escherichia coli) OPCA04413-100UG - FIMH Recombinant Protein (Escherichia coli) |
| OPCA04413-100UG | Aviva Systems Biology | 100ug | EUR 792 |
- FIMH Recombinant Protein (Escherichia coli)
|
) OPCA04413-500UG - FIMH Recombinant Protein (Escherichia coli) OPCA04413-500UG - FIMH Recombinant Protein (Escherichia coli) |
| OPCA04413-500UG | Aviva Systems Biology | 500ug | EUR 1668 |
- FIMH Recombinant Protein (Escherichia coli)
|
 Recombinant E.coli Protein FimH Protein, His, Yeast-1mg Recombinant E.coli Protein FimH Protein, His, Yeast-1mg |
| QP7372-ye-1mg | EnQuireBio | 1mg | EUR 3296.4 |
 Recombinant E.coli Protein FimH Protein, His, Yeast-10ug Recombinant E.coli Protein FimH Protein, His, Yeast-10ug |
| QP7372-ye-10ug | EnQuireBio | 10ug | EUR 434.4 |
 Recombinant E.coli Protein FimH Protein, His, Yeast-50ug Recombinant E.coli Protein FimH Protein, His, Yeast-50ug |
| QP7372-ye-50ug | EnQuireBio | 50ug | EUR 576 |
 Recombinant E.coli Protein FimH Protein, His, Yeast-100ug Recombinant E.coli Protein FimH Protein, His, Yeast-100ug |
| QP7372-ye-100ug | EnQuireBio | 100ug | EUR 945.6 |
 Recombinant E.coli Protein FimH Protein, His, Yeast-200ug Recombinant E.coli Protein FimH Protein, His, Yeast-200ug |
| QP7372-ye-200ug | EnQuireBio | 200ug | EUR 1512 |
 Recombinant E.coli Protein FimH Protein, His, Yeast-500ug Recombinant E.coli Protein FimH Protein, His, Yeast-500ug |
| QP7372-ye-500ug | EnQuireBio | 500ug | EUR 2164.8 |
 Recombinant E.coli Protein FimH Protein, His-GST, E.coli-1mg Recombinant E.coli Protein FimH Protein, His-GST, E.coli-1mg |
| QP7372-ec-1mg | EnQuireBio | 1mg | EUR 2763.6 |
 Recombinant E.coli Protein FimH Protein, His-GST, E.coli-10ug Recombinant E.coli Protein FimH Protein, His-GST, E.coli-10ug |
| QP7372-ec-10ug | EnQuireBio | 10ug | EUR 391.2 |
 Recombinant E.coli Protein FimH Protein, His-GST, E.coli-50ug Recombinant E.coli Protein FimH Protein, His-GST, E.coli-50ug |
| QP7372-ec-50ug | EnQuireBio | 50ug | EUR 522 |
 Recombinant E.coli Protein FimH Protein, His-GST, E.coli-100ug Recombinant E.coli Protein FimH Protein, His-GST, E.coli-100ug |
| QP7372-ec-100ug | EnQuireBio | 100ug | EUR 848.4 |
 Recombinant E.coli Protein FimH Protein, His-GST, E.coli-200ug Recombinant E.coli Protein FimH Protein, His-GST, E.coli-200ug |
| QP7372-ec-200ug | EnQuireBio | 200ug | EUR 1338 |
 Recombinant E.coli Protein FimH Protein, His-GST, E.coli-500ug Recombinant E.coli Protein FimH Protein, His-GST, E.coli-500ug |
| QP7372-ec-500ug | EnQuireBio | 500ug | EUR 1816.8 |
) Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| 1-CSB-YP362349ENV | Cusabio | - Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
| - 100ug
- 10ug
- 1MG
- 200ug
- 500ug
- 50ug
|
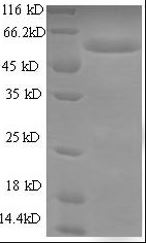
- MW: 31.1 kDa
- Buffer composition: Tris-based buffer with 50% glycerol.
|
Description: Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin(fimH) expressed in Yeast |
| Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| 1-CSB-YP362349ENVe1 | Cusabio | - Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
| - 100ug
- 10ug
- 1MG
- 200ug
- 500ug
- 50ug
|
- MW: 29.1 kDa
- Buffer composition: Tris-based buffer with 50% glycerol.
|
Description: Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin(fimH) expressed in Yeast |
| Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| 1-CSB-EP362349ENV | Cusabio | - Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
| - 100ug
- 10ug
- 1MG
- 200ug
- 500ug
- 50ug
|
- MW: 59.1 kDa
- Buffer composition: Tris-based buffer with 50% glycerol.
|
Description: Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin(fimH) expressed in E.coli |
| Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| 1-CSB-EP362349ENVa0 | Cusabio | - Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
| - 100ug
- 10ug
- 1MG
- 200ug
- 500ug
- 50ug
|
- MW: 33.1kDa
- Buffer composition: Tris-based buffer with 50% glycerol.
|
Description: Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin(fimH) expressed in E.coli |
| Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| 1-CSB-EP362349ENVc7 | Cusabio | - Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
- Ask for price
| - 100ug
- 10ug
- 1MG
- 200ug
- 500ug
- 50ug
|
- MW: 29.8 kDa
- Buffer composition: Tris-based buffer with 50% glycerol.
|
Description: Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin(fimH) expressed in E.coli |
) Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| CSB-YP362349ENV | Cusabio | 6394 mg | Ask for price |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| CSB-YP362349ENVc7 | Cusabio | 7334 mg | Ask for price |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| CSB-YP362349ENVe1 | Cusabio | 6395 mg | Ask for price |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| CSB-EP362349ENV | Cusabio | 3401 mg | Ask for price |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| CSB-EP362349ENVa0 | Cusabio | 3402 mg | Ask for price |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| CSB-EP362349ENVc7 | Cusabio | 3403 mg | Ask for price |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| RPC20134-100ug | Biomatik Corporation | 100ug | EUR 721.6 |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| RPC20134-1mg | Biomatik Corporation | 1mg | EUR 2884.7 |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| RPC20134-20ug | Biomatik Corporation | 20ug | EUR 448.1 |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| RPC25275-100ug | Biomatik Corporation | 100ug | EUR 817.3 |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| RPC25275-1mg | Biomatik Corporation | 1mg | EUR 3461.9 |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| RPC25275-20ug | Biomatik Corporation | 20ug | EUR 479.6 |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| RPC25688-100ug | Biomatik Corporation | 100ug | EUR 721.6 |
| Recombinant Escherichia coli Type 1 fimbrin D-mannose specific adhesin (fimH) |
| RPC25688-1mg | Biomatik Corporation | 1mg | EUR 2884.7 |


 OPCA02151-1MG - fimH Recombinant Protein
OPCA02151-1MG - fimH Recombinant Protein OPCA02951-1MG - fimH Recombinant Protein
OPCA02951-1MG - fimH Recombinant Protein OPCA02151-20UG - fimH Recombinant Protein
OPCA02151-20UG - fimH Recombinant Protein OPCA02151-100UG - fimH Recombinant Protein
OPCA02151-100UG - fimH Recombinant Protein OPCA02951-100UG - fimH Recombinant Protein
OPCA02951-100UG - fimH Recombinant Protein